Las nanopartículas de óxido de zinc mejoran la toxicidad renal inducida por dimetilnitrosamina en ratas
Mar 27, 2022
Contacto: Tina Xiang Correo electrónico:tina.xiang@wecistanche.com
Resumen
dimetilnitrosamina(DMN) es un carcinógeno establecido. Es tóxico para varios órganos, a saber, elhígado, riñón, pulmones y sistema inmunológico. Varios fármacos se han utilizado en el pasado para modular su toxicidad utilizando modelos animales de experimentación. El presente estudio fue diseñado para investigar el efecto de las nanopartículas de óxido de zinc (ZnONP) sobre la toxicidad renal causada por DMN en ratas de laboratorio. Dado que los mecanismos oxidativos están principalmente implicados en su toxicidad, el estudio propuesto se centra en la mejora deestrés oxidativorespuesta por ZnONPs, en su caso. Los presentes resultados muestran que la administración de ZnONP (50 mg/kg de peso corporal/rata) a ratas tratadas con DMN (2 ul/100 g de peso corporal/rata) reduce la concentración de malonaldehído, H, O y NO en el riñón. Sin embargo, la concentración de glutatión reducido (GSH) aumentó después del tratamiento con ZnONP. Los resultados sobre la glutatión S-transferasa y la glutatión peroxidasa favorecieron sus efectos antioxidantes. Estos resultados están respaldados por la recuperación del daño oxidativo del ADN y cambios histopatológicos menos pronunciados en el riñón. Se plantea la hipótesis de que las ZnONP podrían ser tóxicas para el tejido renal; sin embargo, su fuerte potencial terapéutico/antioxidante ayuda a mejorar el daño inducido por DMN.toxicidad renalen ratas
Palabras clave:Dimetilnitrosamina. Riñón. Nanopartículas de óxido de zinc · Estrés oxidativo. Histopatología

Haga clic aquí para obtener más información sobre los efectos de la cistanche
Introducción
La dimetilnitrosamina (DMN) es un carcinógeno establecido [2]. Se ha confirmado que el sitio preferencial de su biotransformación es el hígado. Sin embargo, los órganos, a saber, elriñón, y los pulmones, también participan en su metabolismo, aunque en menor grado [25]. Magee y Barnes [29] demostraron por primera vez que una sola dosis de DMN podía inducir tumores renales. Estudios posteriores atribuyeron el cáncer renal inducido por DMN a especies reactivas de oxígeno (ROS) yestrés oxidativo [3, 32].
Se realizaron ciertos estudios para modular la toxicidad de DMN en modelos animales adecuados utilizando varios fármacos y antioxidantes. Hamza et al. [20] estudiaron los efectos terapéuticos del ácido -lipoico (ALA) contra la inducción de DMNtoxicidad renalen ratas Rana y Kumar [40] informaron que tanto el cadmio como la metalotioneína de zinc inhibían la peroxidación lipídica (LPO) en el riñón de ratas tratadas con DMN. Anteriormente se sabía que el zinc metálico desempeña un papel importante como factor de transcripción y defensa antioxidante en la prevención de la toxicidad por DMN. Los canales de zinc crean un equilibrio entre la supervivencia celular y la muerte celular mediante el control de los movimientos de zinc libres e intracelulares [8]. Por lo tanto, el zinc en el pasado se consideró como un agente adecuado para prevenir la toxicidad de varios xenobióticos, es decir, tetracloruro de carbono |41, alcohol etílico [62] y diclorodifeniltricloroetano [12].
Los recientes avances en nanomedicina han empleado nanopartículas en el tratamiento y diagnóstico de varias enfermedades. En este contexto, varias nanopartículas están siendo sintetizadas y probadas por su toxicidad [22]. Las nanopartículas de óxido de zinc (ZnONP) se han considerado potentes agentes terapéuticos debido a su biodisponibilidad, biocompatibilidad y alta solubilidad. Posee la capacidad de regular el ciclo celular y la homeostasis celular [56]. La Administración de Alimentos y Medicamentos (FDA) también ha aprobado nanopartículas de óxido de zinc para la terapia contra el cáncer[47. Puede causar toxicidad selectiva hacia las células cancerosas por desequilibrio de la actividad de la proteína dependiente de zinc (Vinderall y Mitjans, 2015). Rasmussen et al. [44] planteó la hipótesis de que las ZnONP pueden destruir las células cancerosas mediante la inducción deestrés oxidativo. Por lo tanto, las ZnONP han surgido como plataformas de nano teranóstica contra varias enfermedades, especialmente aquellas causadas por el estrés oxidativo. No obstante, varios laboratorios han publicado informes que muestran su toxicidad en órganos y líneas celulares específicos [11,26].
Por lo tanto, parece haber razones suficientes para examinar más a fondo los efectos antioxidantes de las ZnONP que se manifiestan en la configuración experimental adecuada. Con esta perspectiva, recientemente se realizó en nuestro laboratorio un estudio sobre los efectos protectores de las ZnONPs frente a la hepatotoxicidad inducida por DMN en ratas Wistar macho |43]. Para ampliar este estudio, se hizo un esfuerzo por evaluar los efectos protectores de las ZnONP, si los hubiera, sobre la toxicidad renal inducida por DMN en ratas. Además, también se ha estudiado simultáneamente la toxicidad renal de las ZnONP.

Materiales y métodos
Sustancias químicas y reactivos
Las nanopartículas de óxido de zinc se adquirieron de Sigma Chemical Co. Missouri (EE. UU.). Según el fabricante, las nanopartículas contenían aproximadamente un 80 % de base de zinc, un 100 % de pureza y<100 nm="" size="" with="" a="" surface="" area="" of="" 15-25="">
Dimetilnitrosamina, ácido tiobarbitúrico, ácido 5'-5'-ditiobis-2-nitrobenzoico,1-cloro-2,4-dinitrobenceno, glutatión reductasa, glutatión y N- (1-naftil)etilendiamina diclorhidrato (NEDA) también se adquirieron de Sigma Chemical Co. (EE.UU.). Todos los demás reactivos de la más alta pureza se obtuvieron de High Media (Mumbai).
Caracterización de Nanopartículas de Óxido de Zinc
Las ZnONP se caracterizaron utilizando una batería de métodos como se describió anteriormente [43]. En resumen, el tamaño y la forma de las ZnONP se observaron a través de un microscopio electrónico de transmisión en el Centro de instrumentos analíticos sofisticados de la Universidad de Punjab, Chandigarh (India). Las observaciones de microscopía electrónica de barrido y el análisis de rayos X de dispersión de energía (EDAX) se realizaron en el Departamento de Física de la Universidad Choudhary Charan Singh, Meerut (India). Los análisis de tamaño, distribución y potencial zeta y análisis XRD de ZnONP se realizaron en el Instituto Indio de Tecnología, Roorkee (India).
Mantenimiento de Animales y Protocolo Experimental
Se solicitó aprobación previa del Comité Institucional de Ética Animal para realizar las presentes investigaciones. Los experimentos se realizaron en ratas Wistar macho (150 ± 25 g), obtenidas de las instalaciones para animales de Jamia Hamdard, Delhi. Las ratas se mantuvieron en condiciones estándar de laboratorio (temperatura ambiente, 25 ± 5 grados; humedad relativa, 50 más 10 por ciento; y un ciclo de oscuridad/luz de 12- horas). Cada rata se alojó individualmente en una jaula de polipropileno y se le ofreció comida comercial (Golden Feeds, Delhi) y agua del grifo ad libitum.
Después de la aclimatación a las condiciones de laboratorio durante 2 semanas, las ratas se dividieron al azar en cuatro grupos, cada uno con cinco ratas. A las ratas del grupo A se les inyectó DMN (2 μL/100 g de peso corporal) en solución salina por vía intraperitoneal (ip) en días alternos durante 15 días como se describió anteriormente [43]. Las ratas del grupo B se trataron como las ratas del grupo A y posteriormente se les administró un NOEL predeterminado de ZnONP (50 mg/kg) en días alternos durante 30 días. Las ratas del grupo C se trataron con ZnONP solo como las ratas del grupo B. A las ratas del grupo D se les inyectó (ip) solución salina (2 ul/100 g de peso corporal) solo en días alternos durante 45 días y se trataron como controles.
Después de 45 días, las ratas pasaron hambre durante la noche y sus muestras de orina se recolectaron a la mañana siguiente a través de jaulas metabólicas. A continuación, las ratas se sacrificaron mediante anestesia ligera con éter. losriñonesfueron cuidadosamente removidos y procesados para la estimación de especies reactivas, a saber, malondialdehído, óxido nítrico y peróxido de hidrógeno. El estrés oxidativo se determinó a través de parámetros estándar. es decir, glutatión reducido (GSH), glutatión S-transferasa y glutatión peroxidasa como se describe a continuación.
Creatinina
La creatinina en las muestras de orina se determinó siguiendo el método de Toro y Acker-man (1975), utilizando un kit comercial adquirido de M/S Span Diagnostics, Surat (Gujarat, India)
estrés oxidativo
Malondialdehído (MDA)
La MDA en el tejido renal se determinó usando ácido tiobarbitúrico siguiendo el método de Jordan y Schenkman, [24]. Se registró la absorbancia a 532 nm usando un espectrofotómetro (Systronics, India). Se usó 1,1,3,3-tetrametoxipropano (Sigma) como estándar. La proteína se determinó siguiendo el método de Lowry et al.[27]. Se usó albúmina de suero bovino (Sigma) como estándar.
Peróxido de hidrógeno (H202)
Se prepararon homogeneizados de riñón en sacarosa 0, 25 M. El H2O2 se midió empleando el método del tiocianato férrico descrito por Thurman et al. [52]. La absorbancia se registró a 480 nm utilizando un espectrofotómetro (Systronics, India).
Óxido Nítrico (NO)
El NO en las muestras de riñón se estimó mediante el reactivo de Griess siguiendo el método sugerido por Cortas y Wakid [6]. La absorbancia se registró a 550 nm usando un espectrofotómetro (Systronics, India).
GSH/sulfhidrilos no proteicos (NPSH)
El reactivo de Ellman se utilizó para determinar el glutatión reducido en muestras de riñón [10]. La absorbancia se registró a 412 nm usando un espectrofotómetro (Systronics, India).
Glutatión S-transferasa
La glutatión S-transferasa se analizó usando 1-cloro-2,4-dinitrobenceno (CDNB) que se conjugó con glutatión. La absorbancia se registró a 340 nm [19].
Peróxido de glutation
La enzima se ensayó siguiendo el método de Paglia y Valentine [37]. El disulfuro de glutatión (GSSG) producido como resultado de la glutatión peroxidasa se reduce por un exceso de glutatión reductasa. La conversión de GSSG a GSH se controló a 340 nm utilizando un espectrofotómetro (Systronics, India).
metalotioneína
La metalotioneína en muestras de riñón se analizó siguiendo el método de saturación de cadmio [36], utilizando un espectrofotómetro de absorción atómica (EC, Hyderabad, India).
8-Hidroxi-2'-desoxiguanosina (8-OHdG)
La muestra de orina de cada rata se recogió en un vial esterilizado a través de una jaula metabólica. Estas muestras se almacenaron a -80 grados hasta análisis posteriores. Se usó la técnica ELISA competitiva para la estimación de 8-OHdG utilizando un kit comercial adquirido de Bioassay Technology Laboratory (China). La absorbancia se registró a 450 nm utilizando un lector de microplacas (EC, Hyderabad, India).
Histopatología
Se fijaron pequeños trozos de riñón en formaldehído neutro al 10 por ciento, se deshidrataron, se aclararon y se incluyeron en cera de parafina. Secciones de cinco μm de espesor se tiñeron con hematoxilina y eosina y se examinaron bajo un microscopio de investigación (Nikon, Japón).
Análisis estadístico
Se empleó la prueba t de Student para hacer comparaciones intergrupales entre diferentes grupos. Diferencias entre grupos con valor p<0.05 were="" considered="" significant.="" spss="" software="" version="" 2.0="" was="" used="" for="" inter-group="">

Resultados
Caracterización de ZnONP
La forma, el tamaño, la estructura y la composición eléctrica de las ZnONP se determinaron aplicando métodos estándar. Los resultados muestran que el diámetro promedio de ZnONPs fue<100 nm="" (fig.1a).="" sem="" observations="" showed="" agglomeration="" of="" nps="" (fig.1b).="" the="" electrical="" components="" of="" the="" znonps="" were="" determined="" through="" edax.="" the="" xrd="" pattern="" of="" znonps="" showed="" a="" hexagonal="" structure="" when="" compared="" with="" the="" standard="" data="" (jspds,="" 00-001-1136)="" published="" elsewhere="" [43].="" the="" zeta="" potential="" of="" the="" nanoparticles="" was="" recorded="" to="" be="" 18.9mv(fig.2).="" the="" intensity-weighed="" particle="" size="" distribution="" of="" znonps="" has="" been="" shown="" in="">

 is presented in this fgure")
 of ZnONPs is shown in this fgure. It confrms the size distribution of the particles")
Función renal
Una mayor concentración de creatinina urinaria mostró insuficiencia renal en ratas tratadas con DMN. El tratamiento posterior de ratas tratadas con DMN con ZnONP disminuyó los valores de creatinina. Las ratas tratadas con ZnONP solo también mostraron valores más altos de creatinina que las ratas de control (Tabla 1).
MDA, H2O2 y NO
El malondialdehído (MDA), es el producto de la LPO. aumentó en el riñón de ratas tratadas con DMN. La administración de ZnONP a ratas tratadas con DMN redujo su concentración en el tejido renal. Sin embargo, la concentración de MDA fue mayor en el riñón de las ratas tratadas con ZnONP que en las ratas de control (Tabla 1).
Los valores más altos de NO en el tejido renal de las ratas tratadas con DMN respaldaron los resultados del malondialdehído. La terapia con ZnONP ofrecida a ratas tratadas con DMN disminuyó la concentración de NO en el riñón. Una comparación de los valores de NO obtenidos en el riñón de ZnONP y ratas de control mostró valores más altos aunque insignificantes en el riñón de ratas tratadas con ZnONP (Tabla 1).
Los resultados del peróxido de hidrógeno también mostraron valores más altos en el riñón de ratas tratadas con DMN. Los valores promedio de H, O, en el riñón de las ratas tratadas con DMN más ZnONP fueron más bajos que las ratas tratadas con DMN (Tabla 1). En conjunto, todos estos resultados sugieren un papel antiperoxidativo y antinitrosativo de las ZnONP.
GSH
En el riñón de ratas tratadas con DMN, se observó una disminución significativa de los valores de GSH. Su estado mejoró después de la administración de ZnONPs a ratas tratadas con DMN. Estas observaciones muestran que las ZnONP ofrecen protección antioxidante contra la toxicidad renal inducida por DMN. El tratamiento de ratas con ZnONP mejoró la concentración renal de GSH (Tabla 1).
Glutatión S-transferasa y glutatión peroxidasa
Los resultados sobre GSH fueron respaldados por las observaciones sobre la glutatión S-transferasa. La actividad enzimática disminuyó en el riñón de las ratas tratadas con DMN, la suplementación de ZnONP a las ratas tratadas con DMN restableció su actividad cerca de los valores de control (Tabla 1). La actividad de la glutatión peroxidasa también disminuyó en el riñón de las ratas tratadas con DMN. Sin embargo, aumentó en el riñón de ratas tratadas con DMN y ZnONP (Tabla 1).

8-OHG
Los presentes resultados sobre 8-OHdG mostraron un mayor daño oxidativo del ADN en el riñón de ratas tratadas con DMN. La suplementación con ZnONP en ratas tratadas con DMN inhibió significativamente este daño hasta cierto punto. Sin embargo, el tratamiento con ZnONP solo también podría inducir daños en el ADN en el riñón de ratas (Fig. 4).
")
metalotioneína
Los resultados de la concentración renal de metalotioneína (MT) sugirieron que la inducción de MT disminuyó en el riñón de las ratas tratadas con DMN. Sin embargo, se registró un aumento del 16 por ciento en MT en el riñón de ratas tratadas con DMN y ZnONP. También se encontró que las ZnONP, solas, son un potente inductor de MT en riñones de rata (Tabla 1).

Histopatología
Además de glomerulonefritis y necrosis tubular proximal, se registró adenocarcinoma en la corteza subcapsular en el riñón de ratas tratadas con DMN. La degeneración epitelial fue notoria en los túbulos proximales y distales, se observaron núcleos de diferentes formas y tamaños en toda la corteza y la médula (Fig. 5A, B, C).
Las observaciones histopatológicas en el riñón de ratas tratadas con DMN y ZnONP indicaron una glomerulonefritis menos grave y una necrosis tubular reducida. Faltaba adenocarcinoma. Sin embargo, se observó formación de tejido neoplásico en algunos lugares de la corteza proximal. Se encontró que el epitelio tubular estaba intacto. Los cambios nucleares fueron insignificantes (Figs. 5D, 6A).
Las observaciones histopatológicas en el riñón de ratas tratadas con ZnONP no mostraron nefritis. Los túbulos proximal y distal estaban bien formados y no mostraban signos de degeneración epitelial. Se encontró que el borde del pincel estaba intacto. Sin embargo, se observó un aumento de la actividad mitótica en la corteza distal. (Fig. 6B, C).
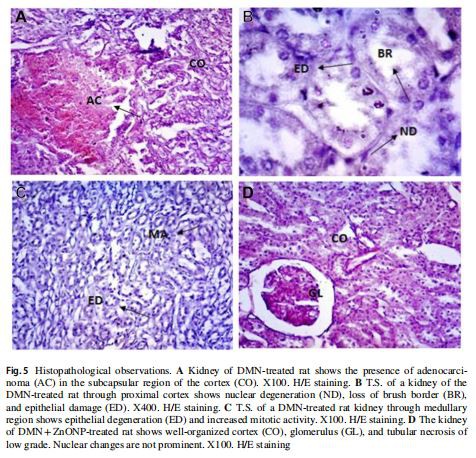
Todos los cambios patológicos descritos anteriormente faltaban en el riñón de las ratas de control. La corteza tubular renal y la médula no mostraban signos de lesión. Se observaron los núcleos normales en la corteza, así como la médula (Fig. 6D).

El presente estudio demuestra que la DMN es igualmente dañina para elriñóncomo es a lahígadoy pulmones El mecanismo de su toxicidad ha sido discutido por algunos trabajadores en el pasado. Se ha establecido ahora quedimetilnitrosaminay otros compuestos nitrosos se metabolizan preferentemente en el hígado; sin embargo, el riñón sí participa en su biodegradación. DMN es metabolizado por CYP2E1 que hidroxila un grupo metilo. La hidroximetil nitrosamina resultante es inestable y se descompone en formaldehído que metila el ADN y las proteínas o reacciona con el agua para formar metanol [13]. La formación de especies reactivas de oxígeno (ROS) como el peróxido de hidrógeno (H2O2) y los radicales hidroxilo (OH) contribuye aestrés oxidativoque podría ser uno de los factores clave en la inducción de cambios patológicos, carcinogenicidad, cambios neoplásicos y formación de tumores no solo en el hígado sino también en los riñones y los pulmones ([57]).

La restauración de la función renal sigue siendo un desafío en la lesión renal tóxica. Dado que se descubrió que las ZnONP protegen contra la lesión hepática inducida por DMN en ratas [43], se consideró esencial un estudio similar en los riñones para demostrar el potencial terapéutico de las ZnONP. La primera indicación de un efecto beneficioso de ZnONPs contra la toxicidad de DMN fue exhibida por observaciones sobre la creatinina. Se elevó en las muestras de orina de las ratas tratadas con DMN, pero disminuyó en las ratas tratadas con DMN y ZnONP. El tratamiento con ZnONP solo también aumentó la concentración de creatinina. La creatinina urinaria/sérica elevada es un biomarcador fiable de la función renal [4]. Se asocia con una función glomerular anormal [5]. Ali Noori et al. [35] también informaron que el tratamiento de ratones Balb/c con ZnONP (50-300 mg/kg) aumentó la concentración de creatinina sérica. Lo correlacionaron con degeneración glomerular y tubular. Durante el presente estudio también. encontramos una correlación entre la concentración de creatinina y los cambios morfológicos renales. La morfología tubular y glomerular renal mejorada en ratas tratadas con DMN y ZnONP se correspondió con una disminución en la concentración de creatinina en la orina. Sin embargo, las ZnONP en la concentración actual y el régimen de dosis mostraron unatoxicidad renal.
Varios estudios han demostrado que el metabolismo de DMN genera ROS en elhígadode animales de experimentación que conducen aestrés oxidativo[18].). Sin embargo, muy pocos trabajadores han demostrado que las ERO también son responsables de su toxicidad renal [54]. Los presentes resultados confirman que DMN podría inducir LPO en elriñóntambién. El tratamiento posterior con ZnONP inhibió la generación de ROS. Dawei et al. [7] postularon que las nanopartículas de óxido de zinc poseen la capacidad de disminuir el malondialdehído y aumentar la actividad de las enzimas antioxidantes. Por el contrario, el malondialdehído también aumentó en el riñón de las ratas tratadas con ZnONP. Otros experimentos realizados sobre la toxicidad de las ZnONP también han revelado que eleva la concentración de MDA en el pez cebra [63] y en el hígado humano [46].

Óxidos nítricos, en elriñónde ratas tratadas con DMN, también mostró valores elevados. Disminuyó en el riñón de ratas tratadas con DMN y ZnONP. Estudios anteriores muestran que los donantes de óxido nítrico como el NaNO previenen parcialmente la hepatitis crónica inducida por la dimetilnitrosamina [28]. Las ZnONP podrían haber afectado la toxicidad renal inducida por DMN al modular la NO sintasa. Los inhibidores de la óxido nítrico sintasa como la no-nitro-L-arginina (L-NNA) podrían atenuar los efectos protectores sobre la toxicidad de la DMN expresada por los donantes de óxido nítrico [14]. H, O, es un importante producto metabólico de DMN [38]. Se registraron valores elevados de H, O, en el riñón de ratas tratadas con DMN. Sin embargo, se registró una disminución en las ratas tratadas con DMN y ZnONP. Esta observación sugiere que las ZnONP influyen en el metabolismo de la DMN. Esta influencia podría estar al nivel de CYP2E1. Sin embargo, se necesitan más estudios para confirmar esta presunción.
Aumento significativo en la concentración renal de MDA, H, O y NO correspondido con una depresión significativa de GSH en el riñón de ratas tratadas con DMN. La administración posterior de ZnONP a ratas tratadas con DMN restauró el estado de GSH en el riñón. El tratamiento con ZnONP en ratas normales también elevó los niveles de GSH. Se sabe que el GSH, un antioxidante no enzimático, contrarresta los efectos dañinos de las ROS [42]. Las ZnONP expresan efectos antioxidantes que pueden atribuirse a su potencial antiinflamatorio mediado por la regulación a la baja de la óxido nítrico sintasa inducible (iNOS), la ciclooxigenasa-2 y varias citocinas [34]. Otros trabajadores atribuyen los efectos beneficiosos de las ZnONP a la metalotioneína [23,33]. En un estudio anterior, Rana y Kumar [40] demostraron que la metalotioneína protege contra la toxicidad de la DMN. Según Durham y Palmiter [9], parece haber una gran posibilidad de que, al liberarse, el zinc actúe como un mensajero compensatorio del estrés oxidativo, estimulando un factor en la región potenciadora del gen MT. La transcripción mejorada de estos genes podría explicar los niveles elevados de Zn-MT en las células estresadas por oxidantes. Los genes para MT y GSH determinan la protección de los inductores de MT [16].
Los presentes resultados muestran que DMN inhibe MT en el riñón en comparación con su concentración en riñones de rata normales. La concentración de MT aumentó en el riñón de ratas tratadas con DMN y ZnONP. La administración de ZnONP solo aumentó significativamente la concentración de MT en el tejido renal. Estos resultados sugieren que las ZnONP también son fuertes inductores de MT Informes anteriores muestran que el zinc es el inductor potencial de MT[30]. MT intercambia zinc relativamente rápido en reacciones intramoleculares e intermoleculares con otros grupos de zinc/azufre a pesar de una estabilidad termodinámica relativamente alta [31].
Se sabe que la DMN afecta la actividad de la glutatión S-transferasa (GST) en el hígado [1,49]. Sin embargo, no se conocen sus efectos sobre las glutatión S-transferasas renales. Las presentes investigaciones mostraron que DMN aumentó la expresión y estimuló la actividad de GST en el riñón. Aniya y Anders [1] informaron que la administración de DMN disminuyó la GST hepática pero la aumentó en el suero. Esta elevación va acompañada de un aumento de la actividad sérica de GPT (SGPT) y de las concentraciones de bilirrubina sérica. Un estudio previo de nuestro laboratorio también ha confirmado la elevación de las transaminasas séricas en ratas tratadas con DMN [43]. El tratamiento de ratas con ZnONP en ratas normales aumentó la actividad de GST en el riñón pero la disminuyó en el riñón de las ratas tratadas con DMN y ZnONP. Sin embargo, no se registró ningún aumento en la concentración renal de GSH. GST y GSH juegan un papel importante en la desintoxicación de mutágenos y carcinógenos [48]. Además, GST puede reducir la unión covalente de epóxidos de carcinógenos como DMN[17].
Muchos trabajadores están de acuerdo en que los efectos protectores de las ZnONP contra el daño inducido químicamente en el hígado/riñón se manifiestan a través de su potencial antioxidante y la prevención de la mutagenicidad y carcinogenicidad mediada por ROS [51]. El tratamiento con DMN en ratas afecta una serie de enzimas antioxidantes, a saber, superóxido dismutasa, catalasa y glutatión peroxidasa. El tratamiento posterior con ZnONP en ratas tratadas con DMN aumentó la actividad de la glutatión peroxidasa en comparación con las ratas de control, lo que indica su mayor capacidad para eliminar H, O , e hidroperóxidos lipídicos [63]. La mejora morfológica en el riñón de ratas tratadas con DMN, manifestada por ZnONP, apoyó las observaciones anteriores. Magee y Barnes [29] confirmaron que DMN podría inducir tumores renales en ratas. Hard y Butler [21] estudiaron la morfogénesis de neoplasias epiteliales inducidas en riñones de rata por DMN. Rio-pelle y Jasmine (1969) clasificaron además los tumores renales inducidos por DMN. Los denominaron islas epiteliales displásicas. Sin embargo, la administración posterior de ZnONP eliminó estos tumores y suprimió otras lesiones morfológicas. La mejora en las enzimas antioxidantes podría haber contribuido a la reparación morfológica en el riñón.
La mayoría de las observaciones discutidas anteriormente favorecen el potencial protector/antioxidante/anticancerígeno de las ZnONP. El presente informe describe la toxicidad de las ZnONP. Una de las características críticas de las ZnONP es su toxicidad selectiva hacia las células cancerosas en comparación con las células normales [39]. Las ZnONP expresan citotoxicidad debido a su composición específica y propiedades de superficie. Las ZnONP son químicamente más activas, conducen a la formación espontánea de ROS en su superficie y causanestrés oxidativo[60]. La formación de ROS contribuye a la toxicidad celular y la liberación de iones Zn plus de las ZnONP debido a su inestabilidad en el compartimento ácido de los lisosomas. Yu et al. [61] y Fukui et al. [15] también concluyó que la toxicidad de ZnONP surge de los iones Zn² más liberados de ZnONP in vitro e in vivo. Wiseman et al. (2006,2007) reveló que el exceso de Zn2 plus libre (disuelto de ZnONP) resultó en el agotamiento de los grupos sulfhidrilo en la metalotioneína y la reducción de la función mitocondrial que condujo a la muerte celular apoptótica o necrótica. Se puede concluir que la toxicidad de ZnONP puede manifestarse a través de varios mecanismos , a saber, el estrés oxidativo, la inhibición de las enzimas antioxidantes, la disfunción mitocondrial y la apoptosis. Curiosamente, el tipo de sistema celular tratado con ZnONP, la fuerza del estrés oxidativo y el entorno intercelular/intracelular existente son factores importantes que determinarán las ZnONP. toxicidad.

Conclusión
En conclusión, el presente estudio sugiere que las ZnONP poseen la eficacia terapéutica potencial para eliminar ROS, inducir GSH y enzimas dependientes de GSH, estimular la síntesis de metalotioneína y reducir el daño oxidativo del ADN. Estos mecanismos, al ser interdependientes, crean un entorno protector contra la toxicidad de las células renales inducida por DMN. Sin embargo, se encontró que las ZnONP son moderadamente tóxicas parariñones. El régimen de dosis debe considerarse como un factor importante en sus efectos protectores.
abreviaturas
DMN: dimetilnitrosamina
ZnONP: nanopartículas de óxido de zinc
NEDA: diclorhidrato de N-(1-naftil)etilendiamina
IP: Intraperitoneal
Zn-MT: metalotioneína de zinc
H2O2: Peróxido de hidrógeno
NO: óxido nítrico
MDA: malondialdehído
GSH: glutatión reducido
ROS: especies reactivas de oxígeno
CDNB: 1-cloro-2,4-dinitrobenceno
8-OHdG: 8-Hidroxi-2'-desoxiguanosina
DA: Adenocarcinoma
CO: Corteza
ND: Degeneración nuclear
BR: Borde de pincel
ED: daño/degeneración epitelial
GL: Glomérulo
MA: actividad mitótica
NPL: neoplasia
NPR: Proliferación nuclear
BC: células binucleadas
EP: revestimiento epitelial
PCT: túbulo contorneado proximal
GL: glomérulos
TEM: microscopio electrónico de transmisión
SEM: microscopio electrónico de barrido
XRD: difracción de rayos X
JSPDS: el comité conjunto sobre estándares de difracción de polvo
EDAX: rayos X de dispersión de energía
Referencias
1. Aniya, Y. y Anders, MW (1985). Alteración de las glutatión S-transferasas hepáticas y liberadas en el suero después del tratamiento con bromobenceno, tetracloruro de carbono o N-nitrosodimetilamina. Farmacología bioquímica, 34, 4239–4244.2. ATSDR, (1989). Perfiles toxicológicos de N-nitrosometilamina. Agencia de Sustancias Tóxicas y Registro de Enfermedades. Atlanta, GA: Departamento de Salud y Servicios Humanos de EE. UU., Servicio de Salud Pública. NAC: 62–75 (9).
3. Bansal, AK, Bansal, M., Soni, G. y Bhatnagar, D. (2005). Modulación del estrés oxidativo inducido por N-nitrosodietilamina (NDEA) por la vitamina E en eritrocitos de rata. Toxicología humana y experimental, 24, 297–302.
4. Bennett, WM (1996). Mecanismos de nefrotoxicidad aguda y crónica por fármacos inmunosupresores. Insuficiencia renal, 18, 453–460.
5. Bishop, LM, Fody, PE y Schoe, HL (2005). Química Clínica. Principios, procedimientos, correlaciones. 5ª ed. Lippincott Williams & Wilkins, Filadelfia, págs. 730. ISBN 0781746116.
6. Cortas, NK y Wakid, NW (1990). Determinación de nitrato inorgánico en muestras de suero y orina por un método cinético de reducción de cadmio. Química clínica, 36, 1440–1443.
7. Dawei, AI, Zhisheng, W. y Angu, Z. (2009). Efectos protectores de nano-ZnO en el cultivo primario de células epiteliales intestinales de ratones in vitro contra la lesión oxidativa. Revista Internacional de Nanotecnología, 3, 1–6.
8. Dhawan, DK y Chadha, VD (2010). Zinc: un agente prometedor en la quimioprevención dietética del cáncer. Revista india de investigación médica, 132, 676–682.
9. Durnam, DM y Palmiter, RD (1981). Regulación transcripcional del gen metalotioneína-I por metales pesados. Revista de Química Biológica, 256, 5712–5716.
10. Ellman, GL (1959). Grupos sulfhidrilo tisulares. Archivos de Bioquímica y Biofísica, 82, 70–77.
11. Fazilati, M. (2013). Investigación de las propiedades de toxicidad de las nanopartículas de óxido de zinc sobre las enzimas hepáticas en la rata macho. Revista Europea de Biología Experimental, 3, 97–103.
12. Feaster, JP, Van Middelem, CH y Davis, GK (1972). Interrelación del zinc DDT en el crecimiento y la reproducción en la rata. Revista de Nutrición, 102, 523–528.
13. Frei, E., Kuchenmeister, F., Gliniorz, R., Breuer, A. y Schmezer, P. (2001). La N-nitrosodimetilamina se activa en los microsomas de los hepatocitos a metabolitos reactivos que dañan el ADN de las células no parenquimatosas en el hígado de rata. Cartas de toxicología, 123, 227–234.
14. Fukawa, A., Kabayashi, O., Yamaguchi, M., Uchida, M. y Hosono, A. (2017). La lactoalbúmina derivada de la leche bovina previene la fibrosis hepática inducida por la dimetilnitrosamina a través de la vía del óxido nítrico en ratas. Biociencia, biotecnología y bioquímica, 81, 1941–1947.
15. Fukui, H., Horie, M., Endoh, S., Kato, H., Fujita, K., Nishio, K., Komaba, LK, Maru, J., Miyauhi, A., Nakamura, A. , Kinugasa, S., Yoshida, Y., Hagihara, Y. e Iwahashi, H. (2012). Asociación de la liberación de iones de zinc y el estrés oxidativo inducido por la instilación intratraqueal de nanopartículas de ZnO en pulmón de rata. Interacciones químicas biológicas, 198, 29–37.
16. Garg, Q. y Hart, BA (1997). Efecto de los tioles en la expresión inducida por cadmio en metalotioneína y otros genes de estrés oxidativo en células epiteliales de pulmón de rata. Toxicología, 119, 179–191.
17. Gopalan, P., Jensen, DE y Lotlikar, PD (1992). Conjugación de glutatión de aflatoxina B1–8 sintética y mediada por microsomas, 9-óxido por glutatión S-transferasas purificadas de ratas. Cartas de cáncer, 64, 225–233.
18. Guengerich, FP, Johnson, WW, Ueng, YF, Yamazaki, H. y Shimada, T. (1996). Participación del citocromo P450, glutatión S-transferasa y epóxido hidrolasa en el metabolismo de la aflatoxina B1 y relevancia en el riesgo de cáncer de hígado humano. Perspectivas de salud ambiental, 104, 557–562.
19. Habig, WH, Pabst, MJ y Jakoby, WB (1974). Glutatión S-transferasas. La primera etapa enzimática en la formación de ácido mercaptúrico. Revista de Química Biológica, 249, 7130–7139.
20. Hamza, RZ, Ismail, HA y El-Shenawy, NS (2017). Estrés oxidativo, alteraciones histopatológicas y microscópicas electrónicas inducidas por dimetilnitrosamina en ratones machos renales y el efecto protector del ácido -lipoico. Revista de fisiología y farmacología básica y clínica, 28, 149–158.
21. Duro, GC y Butler, WH (1971). Morfogénesis de neoplasias epiteliales inducidas en riñón de rata por dimetilnitrosamina. Investigación del cáncer, 31, 1496–1505.
22. Hulla, JE, Sahu, SC y Hayes, AW (2015). Nanotecnología: Historia y futuro. Toxicología humana y experimental, 24, 1318–1321.
23. Jing, L., Li, L., Zhao, J., Zhao, J., Sun, Z. y Perg, S. (2015). La sobreexpresión de metalotioneína inducida por zinc previene la toxicidad de la doxorrubicina en los cardiomiocitos mediante la regulación de las peroxirredoxinas. Xenobióticos, 1, 1–11.
24. Jordan, RA y Schenkman, JB (1982). Relación entre la producción de malondialdehído y el consumo de araquidonato durante la peroxidación de lípidos microsomales soportada por NADPH. Farmacología bioquímica, 31, 1393–1400.
25. Knecht, M. (1966). Sobre la localización de N-desmetilasa microsomal en los órganos de la rata. Naturessenschaften, 53, 85.
26. Li, CH, Shen, CC, Cheng, YW, et al. (2012). Biodistribución, eliminación y genotoxicidad de órganos de nanopartículas de óxido de zinc administradas por vía oral en ratones. Nanotoxicología, 6, 746–756.
27. Lowry, OH, Rosenbrough, NJ, Forr, AL y Randall, RJ (1951). Medición de proteínas con el reactivo de fenol de Follin. Revista de Química Biológica, 193, 265–275.
28. Lukivskaya, O., Lis, R., Zwierz, K. y Buko, V. (2004). Efecto del donante de óxido nítrico y el inhibidor de la óxido nítrico sintasa en el hígado de ratas con hepatitis crónica inducida por dimetilnitrosamina. Revista polaca de farmacología, 56, 599–604.
29. Magee, PN y Barnes, JM (1962). Inducción de tumores renales en rata con dimetilnitrosamina (n-nitrosodimetilamina). Revista de Patología y Bacteriología, 84, 19–31.
30. Maret, W. (2000). La función de la metalotioneína de zinc: un vínculo entre el zinc celular y el estado redox. Revista de Nutrición, 130, 1455–1458.
31. Maret, W., Larsen, KS y Vallee, BL (1997). Dinámica de coordinación de "clusters" biológicos de zinc en metalotioneínas y en el dominio de unión al ADN del factor de transcripción Gal4. Actas de la Academia Nacional de Ciencias de EE. UU., 94, 2233–2237.
32. Mittal, G., Brar, AP y Soni, G. (2006). Impacto de la hipercolesterolemia en la toxicidad de la N-nitrosodietilamina: Efectos bioquímicos e histopatológicos. Informes farmacológicos, 58, 413–419.
33. Mo, R., Jiang, T. y Gu, Z. (2014). Avances recientes en la administración de múltiples fármacos a las células cancerosas mediante liposomas. Nanomedicina, 9, 1117–1120.
34. Nagajyothi, PC, Chan, SJ, Yang, IJ, Sreekanth, TV, Kim, KJ y Shin, HM (2015). Actividades antioxidantes y antiinflamatorias de nanopartículas de óxido de zinc sintetizadas utilizando extracto de raíz de Polygala tenuifolia. Revista de Fotoquímica y Fotobiología B: Biología, 146, 10–17.
35. Noori, A., Karimi, F., Fatahian, S. y Yazdani, F. (2014). Efecto de las nanopartículas de óxido de zinc sobre la función renal en ratones. Revista Internacional de Biociencias, 5, 140–146.
36. Onosaka, S., Tanaka, K., Doi, M. y Okahara, KA (1978). Procedimiento simplificado para la determinación de metalotioneína en tejidos animales. Eisei Kagaku, 24, 128–133.
37. Paglia, DP y Valentine, VM (1967). Estudios de caracterización cuantitativa y cualitativa de la glutatión peroxidasa eritrocitaria. Diario de Laboratorio y Medicina Clínica, 70, 158–169.
38. Pradeep, K., Mohan, CV, Gopichand, K. y Karthikeyan, S. (2007). Efecto de la fístula de Cassia Linn. Extracto de hoja sobre daño hepático inducido por dietilnitrosamina en ratas. Química y Biología, 167, 12–13.
39. Premanathan, M., Karthikeyan, K., Jeyasubramanian, K. y Manivannan, G. (2011). La toxicidad selectiva de las nanopartículas de ZnO hacia bacterias Gram-positivas y células cancerosas por apoptosis a través de la peroxidación lipídica. Nanomedicina, 7, 184–192.
40. Rana, SVS y Kumar, A. (2000). La metalotioneína inducida por cadmio o zinc inhibe la peroxidación lipídica en ratas expuestas a dimetilnitrosamina. Archives of Industrial Hygiene and Toxicology, 51, 279–286.
41. Rana, SVS y Tayal, MK (1981). Influencia del zinc, la vitamina b12 y el glutatión en el hígado de ratas expuestas a tetracloruro de carbono. Salud Industrial, 19, 65–69.
42. Rana, SVS y Kumar, A. (2001). Efecto de la metalotioneína de cadmio y zinc sobre la metahemoglobina y el óxido nítrico en ratas tratadas con dimetilnitrosamina. Revista india de biología experimental, 39, 487–489.
43. Rani, V., Verma, Y., Rana, K. y Rana, SVS (2018). Las nanopartículas de óxido de zinc inhiben la lesión hepática inducida por dimetilnitrosamina en ratas. Interacciones químicas biológicas, 295, 84–92.
44. Rasmussen, JW, Martínez, E., Louka, P. y Wingett, DG (2010). Nanopartículas de óxido de zinc para la destrucción selectiva de células tumorales y potencial para aplicaciones de administración de fármacos. Opinión de expertos sobre la administración de medicamentos, 7, 1063–1077.
45. Riopelle, JL y Jasmin, G. (1969). Naturaleza, clasificación y nomenclatura de los tumores renales inducidos en la rata por dimetilnitrosamina. Revista del Instituto Nacional del Cáncer, 42, 643–662.
46. Sharma, V., Anderson, D. y Dhawan, A. (2012). Las nanopartículas de óxido de zinc inducen daño oxidativo en el ADN y apoptosis mediada por mitocondrias desencadenada por ROS en células hepáticas humanas (HepG2). Apoptosis, 17, 852–870.
47. Shen, C., James, SA, de Jonge, MD, Turney, TW, Wright, PF y Feltis, BN (2013). Relación de la citotoxicidad, los iones de zinc y el oxígeno reactivo en células inmunitarias humanas expuestas a nanopartículas de ZnO. Ciencias toxicológicas, 136, 120–130.
48. Sheweita, SA y Tilmisany, AK (2003). Cáncer y enzimas metabolizadoras de fármacos de fase II. Metabolismo actual de fármacos, 4, 45–58.
49. Sheweita, SA, Mousa, N. y Al-Masry, HM (2008). N-nitrosodimetilamina cambia la expresión de glutatión S-transferasa en el hígado de ratones macho: el papel de los antioxidantes. Revista de toxicología bioquímica y molecular, 22, 389–395.
50. Soheili, S., Moradhaseli, S., Shokouhian, A. y Ghorbani, M. (2013). Efectos histopatológicos de nanopartículas de ZnO en tejidos hepáticos y cardíacos en ratas Wistar. Avances en Bioinvestigación, 4, 83–88.
51. Taccola, L., Rafa, V., Riggio, C., Vittorio, O., Iorio, MC, Vanacore, R., Pietrabissa, A. y Cuschieri, A. (2011). Nanopartículas de óxido de zinc como asesinos selectivos de células en proliferación. Revista Internacional de Medicina Nano, 6, 1129–1140.
52. Thurman, RG, Ley, HG y Scholz, R. (1972). Oxidación de etanol microsomal hepático. Formación de peróxido de hidrógeno y el papel de la catalasa. Revista Europea de Bioquímica, 25, 420–430.
53. Toro, G. y Ackermann, P. (1975). Química clínica práctica, primera ed. Little, Brown and Company, Boston, p.154.





