Flavonoides: ¿un mito o una realidad para la terapia contra el cáncer?
Mar 19, 2022
Para más detalles, póngase en contactotina.xiang@wecistanche.com
Resumen: Los nutracéuticos son moléculas biológicamente activas presentes en los alimentos; pueden tener efectos beneficiosos para la salud, pero no están disponibles en cantidades suficientemente grandes para realizar esta función. Los metabolitos de las plantas, como los polifenoles, se encuentran ampliamente difundidos en el reino vegetal, donde juegan un papel fundamental en el desarrollo de las plantas y en las interacciones con el medio ambiente. Entre estos, los flavonoides son de particular interés ya que tienen efectos significativos en la salud humana. Estudios in vitro y/o in vivo describieron a los flavonoides como nutrientes esenciales para la prevención de diversas enfermedades. Muestran bioactividades amplias y prometedoras para combatir el cáncer,inflamación, infecciones bacterianas, así como para reducir la gravedad de enfermedades neurodegenerativas, cardiovasculares o diabetes. Por lo tanto, no sorprende que el interés por los flavonoides haya aumentado considerablemente en los últimos años. Más de 23,000 publicaciones científicas sobre flavonoides han descrito la potencial actividad anticancerígena de estas moléculas naturales en la última década. Los estudios, in vitro e in vivo, muestran que los flavonoides exhiben propiedades anticancerígenas, y muchos estudios epidemiológicos confirman que la ingesta dietética de flavonoides reduce el riesgo de cáncer. Esta revisión ofrece una idea de los mecanismos de acción de los flavonoides en las células cancerosas.
Palabras clave: flavonoides; cáncer; estrés oxidativo; inflamación; apoptosis/autofagia; metástasis; angiogénesis

1. Introducción
loscáncerLa tasa de mortalidad ha disminuido a lo largo de los años debido a la investigación y la prevención, pero su tasa de incidencia ha aumentado. Varios estudios han destacado el papel de una dieta basada en plantas en la prevención de enfermedades relacionadas con la aparición de tumores [1]. Los beneficios de una dieta basada en plantas podrían derivarse de la presencia de varios componentes bioactivos, como compuestos fenólicos, carotenoides y, en particular, flavonoides, en los vegetales. Estos últimos son considerados indispensables y presentes en diversas aplicaciones nutracéuticas, cosméticas, farmacéuticas, medicinales y cosméticas. Debido a estas aplicaciones, la investigación sobre flavonoides se ha incrementado sustancialmente en los últimos años.
Flavonoides son un subgrupo de metabolitos secundarios pertenecientes a una gran colección de compuestos fenólicos sintetizados por las plantas. Están ampliamente distribuidos entre los organismos fotosintéticos y son abundantes en alimentos y bebidas de origen vegetal (Cuadro 1), donde las composiciones cualitativas y cuantitativas pueden variar considerablemente. La estructura química está compuesta por un esqueleto de 15 átomos de carbono, que contiene dos anillos bencénicos (A y B) unidos al anillo piránico heterocíclico (C)[2]. Los flavonoides se pueden dividir en varios subgrupos: flavonas, flavonoles, flavanonas, flavanonoles, flavanoles o catequinas, antocianinas y chalconas [3]. Esta distinción se deriva de la estructura básica del flavonoide (Figura 1), el anillo de flavona, que representa la parte principal del flavonoide, y el grado de insaturación y oxidación del anillo carbonoso. Además, en las plantas, la aglicona es la estructura básica de los flavonoides; sin embargo, pueden estar presentes éteres metílicos y ésteres acetílicos del grupo alcohol, así como glucósidos formados a través del enlace con un carbohidrato, como L-ramnosa, D-glucosa, glucosa-ramnosa, galactosa o arabinosa [4].

Más de 10,000 moléculas pertenecen al gran grupo de flavonoides [12,13]. Este número aumenta considerablemente si consideramos no solo los productos derivados de los flavonoides y formados durante el procesamiento y almacenamiento de los alimentos, sino también los metabolitos y conjugados producidos en el organismo tras su ingesta. Por lo tanto, las concentraciones de flavonoides y la complejidad estructural y las características fisicoquímicas varían mucho según la fuente y la matriz [14].

Es muy difícil estimar la ingesta dietética de flavonoides debido a su variabilidad cuantitativa y cualitativa en vegetales y frutas, lo que puede dificultar el establecimiento de relaciones epidemiológicas en cuanto a su impacto en la salud y enfermedad humana. Los numerosos estudios sobre absorción y biodisponibilidad presentan en la literatura han sido revisados por diferentes autores [15-17]. Varios factores pueden afectar la biodisponibilidad de los flavonoides, como los pesos moleculares, la glicosilación y la esterificación, lo que provoca un grado de incertidumbre sobre los niveles reales de su biodisponibilidad y absorción en el cuerpo humano [17].
Crozier y colegas [16] y Landete [18] proporcionan una descripción detallada de la conversión metabólica de los flavonoides después de la ingesta dietética. Brevemente, la conversión metabólica de los flavonoides puede tener lugar en el intestino delgado con la liberación de agliconas como resultado de las actividades de las hidrolasas. A este paso le sigue la conversión en el hígado, donde se producen formas conjugadas, es decir, O-glucurónidos, ésteres de sulfato y O-metilésteres de flavonoides. El cuerpo puede tratar estos metabolitos como xenobióticos; por lo tanto, eliminándolos del torrente sanguíneo [16,18]. Los derivados de glucurónidos y sulfatos pueden excretarse más fácilmente a través de la orina y la bilis [18]. En consecuencia, el análisis de plasma puede no proporcionar información valiosa sobre los perfiles de estos metabolitos, mientras que la excreción urinaria presenta una gran variabilidad individual según las clases de flavonoides y la posibilidad de absorción de los metabolitos por los tejidos corporales. Además, los compuestos, no absorbidos por el intestino, pasarán al colon, donde serán modificados estructuralmente por la microflora colónica. Los catabolitos derivados pueden absorberse en el torrente sanguíneo y finalmente excretarse en la orina. Además, los flavonoides pueden modular la composición de la microbiota intestinal aumentando la población de bacterias beneficiosas, por ejemplo, Bifidobacterium y Lactobacillus, e inhibiendo el crecimiento de diferentes patógenos[19]. Tal capacidad de los flavonoides proporciona un importante mecanismo antipolítico.
1.1. Ruta biosintética de flavonoides en plantas
El metabolismo deflavonoidesinvolucra genes ya presentes en las primeras plantas terrestres, hepáticas y musgos [20]. La ruta bioquímica se caracterizó a través del estudio de mutantes con una síntesis alterada de flavonoides presentes en varias especies de plantas [21]. Los precursores clave para la síntesis de flavonoides son la fenilalanina y el malonil-CoA producidos por la vía del shikimato y el ciclo TCA (ciclo del ácido tricarboxílico). A través de la vía del shikimato, se producen aminoácidos aromáticos en plantas, bacterias y hongos. Esta ruta consta de siete reacciones enzimáticas, comenzando desde la reacción entre el fosfoenolpiruvato y el fosfato de eritrosa-4-, hasta la síntesis de corismato, el producto final de la ruta, catalizada por la corismato sintasa. El corismato mutasa reorganiza el corismato para prefenato; este último es el sustrato utilizado para sintetizar fenilalanina [22]. En las plantas, la fenilalanina es el precursor de la 4-cumaroil-CoA, seguida de la actividad de la fenilalanina amoníaco-liasa (PAL) y la 4-cumarato-CoA ligasa. Para iniciar la síntesis de flavonoides, 4-cumaroil-CoA reacciona con malonil-CoA [23] (Figura 2). Estas enzimas se localizan en el lado citosólico del retículo endoplásmico (ER), como lo sugieren los experimentos de localización inmune, y se recuperan en la fracción soluble de los extractos celulares. Además, las enzimas se asocian entre sí mediante interacciones proteína-proteína en la superficie del retículo endoplásmico (RE); formando así un complejo [21,24,25]. Los datos sobre la colocalización de algunas enzimas en el tonoplasto y el núcleo han sugerido un comportamiento dinámico del complejo biosintético. Esto favorecería tanto la canalización como el desplazamiento de los productos finales para satisfacer las necesidades fisiológicas de la célula [24]. Los compuestos se dirigen a las vacuolas como orgánulos de almacenamiento (es decir, antocianinas, flavonol y glucósidos de flavona) oa las paredes celulares [21. Sin embargo, es esencial señalar que, bajo ciertas condiciones fisiológicas, las células vegetales pueden removilizar los flavonoides de los depósitos de vacuolas, por lo que el transporte a través del tonoplasto no es unidireccional [25]. Además de las vacuolas y las paredes celulares, los flavonoides se encuentran en el citosol, ER, cloroplastos (es decir, glucósidos de quercetina y kaempferol), núcleo (es decir, isoflavonoides cumestrol y 4',7-dihidroxiflavona en Medicago truncatula) y vesículas pequeñas, así como espacio apoplásico (es decir, flavona, agliconas de flavonol e isoflavonas) [25]. Un sistema de transporte eficiente de flavonoides dentro de las células parece ser la base de su amplia distribución a diferentes compartimentos celulares. Dos sistemas principales parecen estar involucrados en el transporte de flavonoides, uno basado en vesículas de membrana y el otro en un transportador de membrana, que no parece que se excluyan mutuamente [25].
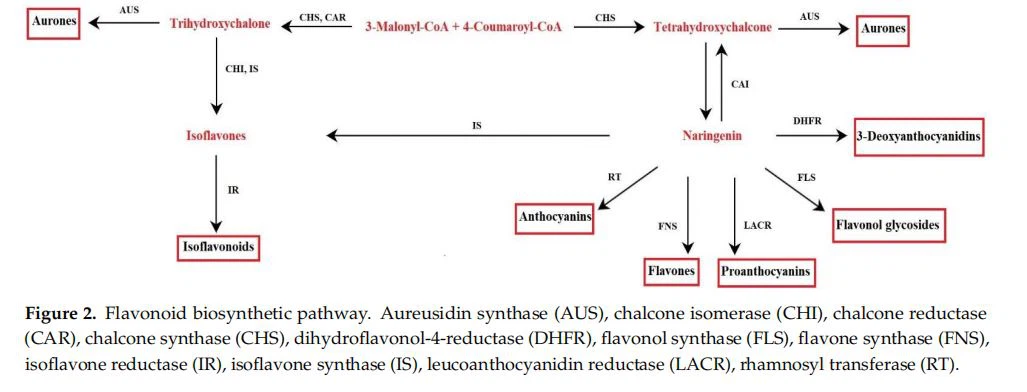
1.2.Papel de los flavonoides en las plantas
La conservación de genes implicados en el metabolismo de los flavonoides durante la evolución de las plantas terrestres es un claro indicio de su papel fundamental en la fisiología de la planta [15]. Los flavonoides son responsables del color y el aroma de las flores, están involucrados en las estrategias reproductivas, protegen las células de la dañina radiación UV (esencial para la vida de las plantas terrestres) y desempeñan un papel en la resistencia a las enfermedades, así como en la asociación simbiótica (es decir, como moléculas señal en la simbiosis planta-microorganismo). Al participar en las respuestas al estrés, protegen a la planta de las duras condiciones ambientales [26-28]. La amplia difusión de los flavonoides sugiere que su actividad antioxidante es una característica sólida para la supervivencia y la aptitud de las plantas terrestres. De hecho, su síntesis mejora después de la exposición de la planta a un estrés severo, ya que su poderosa actividad antioxidante puede contrarrestar los efectos nocivos de las especies reactivas de oxígeno (ROS) [29,30].
1.3. Flavonoides y Biotecnología
Los flavonoides se han asociado con muchas características agronómicas favorables y beneficios para la salud de los humanos, por lo que su ingeniería metabólica es un objetivo importante para la biotecnología vegetal [25]. Las cantidades de flavonoides en las plantas varían según la especie, las condiciones de crecimiento y la etapa de desarrollo. De hecho, incluso si las plantas aromáticas y medicinales son bastante eficientes en la producción de estas moléculas, las plantas cultivadas en el campo no siempre pueden representar una buena fuente de estos metabolitos. Esto se debe a las dificultades en el cultivo de plantas, las variaciones estacionales en la productividad, la producción específica de tejidos/órganos y los problemas relacionados con la purificación. Por estas razones, la producción industrial de polifenoles sería difícil de sostener si las plantas cultivadas en el campo fueran la única fuente de materia prima. Por otro lado, las estructuras altamente complejas y la estereoespecificidad de los flavonoides a menudo hacen que la síntesis química no sea económicamente factible [31]. Las técnicas in vitro pueden representar una herramienta para mejorar la biosíntesis y disponibilidad de flavonoides durante todo el año para superar estos problemas. Se han utilizado diferentes cultivos in vitro de plantas (es decir, callos, cultivos en suspensión celular, cultivos de órganos y raíces peludas) y técnicas de transformación para investigar y mejorar la síntesis de estas importantes moléculas [31-35]. Se han tenido en cuenta varios enfoques, como la selección de líneas de alto rendimiento, la alimentación de precursores y el uso de elicitores [36]. Esta última implica la adición al medio de cultivo de moléculas de síntesis biológica o química, capaces de estimular la acumulación de metabolitos secundarios en la planta como respuesta de defensa a condiciones de estrés [28], disparadas y activadas por elicitores[{{8} },36]. Se han obtenido resultados positivos en diferentes especies [37] y, en esta perspectiva, se puede considerar el uso de elicitores para un futuro desarrollo a escala industrial.
Además, la mejora del conocimiento sobre el papel de los miARN en la regulación de la vía biosintética de los flavonoides permitirá mejorar el metabolismo de estas moléculas. La modulación de los niveles de miARN podría ser una herramienta poderosa tanto para obtener un mejor rendimiento como para la síntesis de las combinaciones deseadas de metabolitos [38].

2. Actividades preventivas del cáncer de los flavonoides
El amplio espectro de acciones biológicas que llevan a cabo los flavonoides depende en gran medida de su característica de ser poderosos antiinflamatorios y antioxidantes que contrarrestan los radicales libres, ligados de manera importante para muchas enfermedades crónico degenerativas (Figura 3). En condiciones patológicas, el aumento de radicales libres daña diferentes tipos de moléculas, como ácidos nucleicos, proteínas y lípidos, y da como resultado el envejecimiento y la muerte celular, pero también promueve la carcinogénesis [39].
![Anticancer potential of flavonoids (from [40] with modifications)](/Content/uploads/2022842169/20220317124220ac179298fe6042fdbf75f6c46c0c4987.png "Anticancer potential of flavonoids (from [40] with modifications)")
2.1. Flavonoides e Inflamación Crónica
Cáncerse considera una enfermedad relacionada con la crónicainflamación[41]. En varias enfermedades inflamatorias, el resultado conduce a la carcinogénesis. En la vía biliar, el colangiocarcinoma produce un infiltrado inflamatorio crónico, debido a la infección por Clonorchis Sinensis [42]. Helicobacter pylori representa una de las principales causas de adenocarcinoma y linfoma del tejido linfoide asociado a la mucosa gástrica [43]. La infección crónica por el virus de la hepatitis B y C puede provocar carcinoma hepatocelular, la tercera causa principal de muerte por cáncer [44]. Finalmente, la infección por el virus del papiloma es una de las principales causas de cáncer de pene y anogenital en humanos. Además, el riesgo de desarrollar cáncer de vejiga puede aumentar después de la esquistosomiasis, al igual que el riesgo de contraer el sarcoma de Kaposi después de la infección por el virus del herpes humano tipo 8. Otras formas de inflamación crónica, además de las generadas por infecciones microbianas, pueden contribuir a la carcinogénesis. Se ha descrito un mayor riesgo de cáncer de páncreas, esófago y vesícula biliar como consecuencia de enfermedades inflamatorias como la metaplasia de Barrett, la esofagitis y la pancreatitis crónica [45, A46]. También se han encontrado posibles asociaciones entre la úlcera de Marjolin y el cáncer de piel [47], el asbesto y el mesotelioma [48], el humo del cigarrillo y el cáncer bronquial [48], el asma crónica y el cáncer de pulmón [49], el liquen plano ulceroso y el carcinoma de células escamosas [ 50], inflamación del prepucio/fimosis y cáncer de pene [51] y entre inflamación pélvica/ovárica y cáncer de ovario [52]. El cáncer de próstata se ha asociado con prostatitis crónica causada por una infección bacteriana persistente o por mecanismos no infecciosos [53]. Por lo tanto, la asociación entre la inflamación crónica y el desarrollo de cáncer parece respaldada por un creciente cuerpo de evidencia.
En este sentido, los flavonoides han demostrado la doble capacidad de reducir la inflamación y la proliferación de células tumorales. La taxifolina, un flavanonol que se encuentra en las coníferas, tiene efectos antiinflamatorios o antiproliferativos. En ratones albinos suizos desafiados con benzopireno, un mutágeno que está frecuentemente presente en el humo del cigarrillo y en los escapes de los automóviles. ejerció una inflamación suprimida a través de la estimulación de la vía de señalización Nrf2 (factor 2 relacionado con el factor nuclear eritroide 2-), que desempeña un papel central en proporcionar resistencia al estrés oxidativo y la inflamación al inhibir NF-kB [54,55]. La crisina es un flavonoide aglicónico con funciones antiinflamatorias. La administración de crisina en ratones desafiados con LPS (lipopolisacárido) redujo el desarrollo de lesiones pulmonares al suprimir la vía de la proteína 3 del receptor similar a la oligomerización del dominio de unión de nucleótidos / proteína de interacción de la enzima 1 / tiorredoxina que requiere inositol [56]. En ratas, previno las complicaciones miocárdicas del estrés oxidativo provocado por la hipercolesterolemia a través de la activación de la óxido nítrico sintasa endotelial y genes diana Nrf2 como SOD (superóxido dismutasa) y catalasa [57]. Además, la crisina inhibió significativamente la proliferación e indujo la apoptosis en células humanas de cáncer de cuello uterino [58] y células de cáncer colorrectal [59] mediante la modulación de varios genes apoptóticos y genes de la vía AKT/MAPK. Estos resultados destacan dos mecanismos distintos a través de los cuales los flavonoides determinan los efectos sobre la inflamación y la proliferación celular: por un lado, activan la vía Nrf2 para inhibir NF-kB y desencadenar el efecto antiinflamatorio; por otro lado, actúan sobre la proliferación celular mediante la modulación de los genes implicados en la apoptosis y la vía AKT/MAPK (protein kinase B/mitogen-activated protein kinase).
2.2. Flavonoides y Estrés Oxidativo
El ambiente intracelular en las células cancerosas tiene un alto nivel de ROS que la célula normal, principalmente peróxido de hidrógeno, debido a un sistema antioxidante que ya no es eficiente. En las células normales, una proporción adecuada de glutatión (GSH/GSSG) convierte el peróxido de hidrógeno en agua. Cuando la proporción de glutatión disminuye, el peróxido de hidrógeno se convierte en radical hidroxilo (OH'), que es un radical muy reactivo que provoca daños en el ADN y mutaciones en los genes supresores de tumores, un evento crítico inicial que desencadena la carcinogénesis [60]. Al menos tres etapas caracterizan el desarrollo del cáncer: iniciación, promoción y progresión.Estrés oxidativoparticipa en todas las fases de este proceso (Figura 4). Durante la fase de iniciación, las ROS pueden dañar el ADN al introducir mutaciones genéticas y alteraciones estructurales en el ADN. En la fase de promoción, las ROS tienen un papel fundamental en el aumento de la proliferación celular o la reducción de la apoptosis celular como consecuencia de la modificación de la expresión génica, la comunicación entre células y las vías de señalización intracelular [61]. Finalmente, el estrés oxidativo contribuye a la progresión del proceso tumoral a través de una mayor mutagénesis en la población celular iniciada [62]. El objetivo terapéutico de muchos medicamentos contra el cáncer es aumentar el nivel ya alto de ROS presente en las células tumorales para desencadenar la cascada de apoptosis [63]. Incluso los flavonoides, aunque reconocidos por su actividad antioxidante, pueden tener actividad prooxidante y, por lo tanto, desencadenar la apoptosis en las células cancerosas.

La naringenina es una flavanona más abundante en la toronja, la mandarina, la naranja, la cáscara de limón cruda y la cáscara de lima cruda. Detuvo el ciclo celular e indujo la apoptosis en varias células tumorales humanas [64,65], y también suprimió la invasividad y el potencial metastásico de las células de cáncer gástrico y las células de carcinoma hepatocelular [66,67]. La naringenina tuvo un efecto prooxidante al reducir las actividades de la glutatión reductasa, la glutatión S-transferasa y la glioxalasa en las células tumorales, lo que a su vez redujo los mecanismos de detoxificación del peróxido de hidrógeno, lo que permitió la acumulación y el aumento de la peroxidación lipídica con el consiguiente daño de la membrana celular. [68]. Curiosamente, un ensayo clínico de fase 1 recientemente concluido ha destacado la seguridad y la farmacocinética de la naringenina [69]. La naringenina, 4h después de la administración de una dosis única de extracto de Citrus sinensis (naranja dulce), fue detectable en plasma a una concentración de 43 μuM.

2.3. Flavonoides y Apoptosis/Autofagia
La búsqueda de terapias contra el cáncer se centra actualmente en la inducción de la apoptosis de las células cancerosas [70]. Desafortunadamente, las células cancerosas pueden evitar la activación de la cascada apoptótica, defendiéndose de la muerte celular. Además, el desarrollo tumoral se ve favorecido por la inducción de resistencia a los fármacos [71]. La modulación de Bcl-2 y otras proteínas, permite que flavonoides, como el yeso, aislado de la especie Vitex agnus-castus, ampliamente utilizada en la medicina tradicional china como agente antiinflamatorio, desencadene la apoptosis modulando Bcl{{ 5}} y otros pro-supervivencia. Esta molécula desencadena la vía intrínseca de la apoptosis mediante la regulación negativa de Bcl-2, Bcl-xL, survivina y la regulación positiva de Bax, como se evidencia en numerosas líneas tumorales de cáncer de vesícula biliar, cáncer de esófago, cáncer de colon, leucemia y glioblastoma [72] . De manera similar, la vitexina es un compuesto flavonoide de origen natural extraído de la hierba china Crataegus pinnatifida que se ha demostrado que reduce la relación Bcl-2/Bax, la liberación de citocromo c de las mitocondrias y en el cáncer de pulmón de células no pequeñas en humanos A549 células, escisión de caspasa-3 [73].
Se observó en el inhibición de la proliferación de una línea de cáncer de ovario metastásico humano (PA-1) ejercida por la quercetina [74], uno de los flavonoides más abundantes en las cebollas y el brócoli.
La autofagia es un proceso catabólico inducido por estrés altamente conservado que regula positivamente el proceso de muerte celular. Varios medicamentos contra el cáncer desencadenaron la autofagia y, por lo tanto, su inducción representa una estrategia potencial para la terapia contra el cáncer J751. El extracto acuoso de Allspice es rico en diferentes tipos de flavonoides. En las células de cáncer de mama, activó la autofagia, in vitro e in vivo, e indujo la muerte celular al suprimir la vía Akt/mammalian target of the rapamycin (mTOR) [76]. De manera similar, en células de cáncer hepático humano SK-HEP-1, el kaempferol indujo autofagia a través de la señalización de Akt y la proteína quinasa activada por monofosfato de adenosina (AMPK), y a través de la regulación a la baja de CDK1/ciclina B condujo a la detención de G2/M [77]. ]. Además, la inducción de autofagia por parte de la genisteína en múltiples tipos de cáncer, como el de mama, próstata y útero, parece ser la base de su efecto antitumoral [78].
2.4. Flavonoides dirigidos a las células madre cancerosas
Las células madre cancerosas (CSC) son una pequeña subpoblación de células en un tumor que se autorrenuevan y son capaces de iniciar y mantener el crecimiento del tumor. Además, las CSC en el cáncer desempeñan un papel fundamental en el inicio, el mantenimiento, la progresión, la resistencia a los medicamentos y la recurrencia o metástasis [79]. La acumulación de evidencia sugiere que los fitoquímicos dietéticos, incluidos los flavonoides, son agentes prometedores para contrarrestar las CSC [80]. Por ejemplo, se ha demostrado que la naringenina inhibe las células madre del cáncer de mama a través del aumento de p53 y del receptor de estrógeno de manera similar a como se encontró con la hesperidina [81].
La apigenina es una flavona común que se encuentra principalmente en la manzanilla, el apio y el perejil. La actividad anticancerígena de la apigenina se ha observado en el glioblastoma (el tumor cerebral primario y agresivo más común). De hecho, Kim y sus colegas [82] demostraron que la apigenina (y la quercetina) pueden interferir con la capacidad de autorrenovación y la invasividad de las células similares a las del glioblastoma a través de la regulación a la baja de la vía de señalización de c-Met. La apigenina aumenta la actividad antineoplásica del cisplatino en las poblaciones de células madre de cáncer de próstata CD44 más [83] y suprime las propiedades similares a las células madre y el potencial tumorigénico de las células de cáncer de mama triple negativas [84]. La inhibición de la capacidad de autorrenovación y la restauración de la radiosensibilidad se han demostrado en las células madre del cáncer oral para la luteolina [85], una flavona que se encuentra en una gran variedad de fuentes dietéticas que incluyen apio, zanahorias, pimientos, aceite de oliva, romero, y orégano. El flavonol quercetina es una molécula de interés médico, ya que posee potencial anticancerígeno [86]. De hecho, la quercetina se dirige a varios tipos de CSC, incluidas las células madre pancreáticas [87], mamarias [88] y gástricas [89].
2.5. Propiedades antiangiogénicas y antimetastásicas de los flavonoides
Los flavonoides juegan un papel interesante como inhibidores de la angiogénesis. La angiogénesis consiste en el desarrollo de nuevos vasos sanguíneos, que es un proceso fundamental para el crecimiento de los tejidos, la cicatrización de heridas y el desarrollo embrionario, pero representa una característica negativa en presencia de un tumor ya que más vasos sanguíneos llevan más nutrientes a las células cancerosas permitiendo para vivir mejor y proliferar. Es un proceso estrechamente controlado por una amplia gama de inductores, como el factor de crecimiento endotelial vascular (VEGF) y moléculas de adhesión, así como por varios inhibidores, como la angiostatina y la trombospondina, y estimulado por muchos factores que contribuyen a la inflamación y el cáncer, lo que indica que la angiogénesis, la inflamación y el cáncer son procesos estrechamente relacionados [90]. En los últimos años, el desarrollo de inhibidores de la angiogénesis ha sido un foco de atención de la investigación contra el cáncer, ya que este proceso descontrolado es un paso fundamental en el crecimiento, la invasión y la metástasis del cáncer. Tras este esfuerzo, la FDA aprobó el uso de numerosos fármacos antiangiogénicos para el tratamiento del cáncer [91]. Se están probando nuevas moléculas capaces de inhibir la angiogénesis tumoral. Wogonin, una flavona O-metilada, un compuesto químico similar a los flavonoides sintetizado por Scutellaria baicalensis, inhibe la angiogénesis inducida por LPS tanto in vitro como in vivo [92]. La genisteína inhibe la angiogénesis al modular la expresión de VEGF, metaloproteasas (MMP) y receptor del factor de crecimiento epidérmico (EGFR) [93]. En las células endoteliales de la vena umbilical humana, estimuladas por VEGF(HUVECs), Kaempferol inhibe la angiogénesis actuando sobre el receptor 2 de VEGF. Este proceso también se lleva a cabo gracias a la regulación a la baja de P13kt/Akt junto con el activado por mitógeno. proteína quinasa (MEK) y las vías ERK [94].
La luteolina (8-C- -D-glucopiranósido), un flavonoide glicosilado de la dieta, reduce la invasión tumoral en MCF tratados con 12-O-tetradecanoilforbol-13-acetato (TPA){{ 7}} células de cáncer de mama, bloqueando la expresión de MMP-9metaloproteinasa e interleucina-8(IL-8)[95]. En las células de cáncer gástrico, la quercetina mostró efectos antimetastásicos a través de la descomposición de la función del activador del plasminógeno de la urocinasa (uPA)/receptor de uPA (uPAR), mediante la modulación de NF-kB, PKC-6, ERK1/2 y AMPK [96]. Recientemente, Yao et al. informaron que en las células de melanoma humano A375, la luteolina inhibe la proliferación, la migración y la invasión al inducir la apoptosis dependiente de la dosis. En el mismo modelo celular, también se observó inhibición de la fosforilación de Akt y PI3K. Los mismos autores han recogido evidencia experimental de que la luteolina permite la sobreexpresión de inhibidores tisulares de la metaloproteinasa (TIMP)-1 y TIMP-2 y reduce la expresión de MMP-2 y MMP{{23} }]. Otros resultados experimentales destacaron que la luteolina redujo significativamente el crecimiento tumoral de las células A375 en un modelo de xenoinjerto de ratón, lo que confirma que la actividad antitumoral se deriva de la regulación a la baja de la expresión de MMP-2 y MMP-9 a través de PI3K/Akt camino [97].
2.6. Flavonoides y diferenciación de células cancerosas
La terapia de diferenciación tiene como objetivo inducir la diferenciación de las células cancerosas; por lo tanto, reduciendo su proliferación [68]. La terapia de diferenciación en comparación con la quimioterapia convencional tiene la ventaja de ser menos tóxica y, por lo tanto, causar menos efectos secundarios al paciente [98]. La quercetina y la pelargonidina inducen la diferenciación en células murinas de melanoma B16-F10 altamente metastásicas mediante un mecanismo que implica la transglutaminasa tipo 2 [99]. El ácido retinoico todo trans (ATRA) tiene un amplio uso clínico en la terapia de diferenciación en pacientes con leucemia promielocítica aguda (APL). Sin embargo, el tratamiento prolongado produce resistencia a los medicamentos y requiere una dosis cada vez más alta [100]. La aparición de fenómenos de resistencia a fármacos necesita el desarrollo de nuevos agentes con mayor actividad inductora de diferenciación. Los flavonoides tienen características interesantes en este sentido. De hecho, son capaces de inducir la diferenciación celular de las células APL. Sin embargo, la estructura de la flavona podría ser crucial para la inducción de la diferenciación celular. De hecho, en las células APL, la quercetina induce su diferenciación en monocitos y la apigenina y la luteolina inducen su diferenciación en granulocitos. Por el contrario, galangina, kaempferol y naringenina no indujeron ninguna diferenciación en las células APL [100].
Recientemente, Moradzadeh et al. [101] informaron que el galato de epigalocatequina (EGCG), un polifenol del té verde, en la diferenciación de granulocitos de células APL HL-60 y NB4, posee un efecto similar al ATRA. En ambas líneas celulares, EGCG redujo la expresión de histona desacetilasa 1. Además, en células NB4, EGCG también redujo la expresión de un marcador clínico relevante PML-RARo. La diferenciación celular fue inducida por wogonina, en la línea celular K562, un modelo celular de leucemia mieloide crónica primaria (LMC). Se observó el mismo resultado en LMC primaria derivada de pacientes que era sensible y resistente a imatinib. En estas células también se observó una regulación al alza del factor de transcripción GATA-1 y una mayor unión entre GATA-1 y el coactivador transcripcional FOG-1 [102]. Varias observaciones brindan evidencia para respaldar la aplicación potencial de los flavonoides en el tratamiento de pacientes con diferentes tipos de cáncer. En células tumorales aisladas de varios tumores sólidos, como melanoma maligno, cáncer de mama, glioma y hepatoma, se ha demostrado la diferenciación inducida por el tratamiento con flavonoides [103]. Específicamente, en las células madre del cáncer de mama, se ha observado la diferenciación celular inducida por la genisteína [78,93] y un flavonoide aislado del regaliz (Glycyrrhiza sp.), la isoliquiritigenina [104].
En el tratamiento de células APL NB4, con dihidromiricetina (DMY), un dihidroflavonol extraído de Ampelopsis sp., se observó que este sinergizaba con ATRA, para promover la diferenciación celular [105]. La fosforilación de p38 MAPK inducida por ATRA activa STAT1, y STAT1 desempeña un papel clave en la diferenciación terminal de las células mieloides a través de la regulación de proteínas del ciclo celular y factores de transcripción mieloides específicos. La diferenciación mejorada por DMY, cuando se combinó con ATRA, dependía de la mayor activación de la vía de señalización p38MAPK/STAT1. Curiosamente, DMY solo no pudo activar la diferenciación y redujo la fosforilación de p38 MAPK con la consiguiente reducción en la actividad de STAT1 [105]. Este comportamiento inesperadamente diferente, en la activación de la vía, sugiere que no es posible predecir el efecto biológico derivado de la combinación de un flavonoide genérico con un fármaco convencional basándose simplemente en el conocimiento de su mecanismo de acción estudiado en tratamientos individuales, ya que puede que no sea lo mismo. Por lo tanto, todos los flavonoides podrían ser posibles potenciadores de la diferenciación en combinación con fármacos convencionales.
2.7. Flavonoides para mejorar la sensibilidad a la quimioterapia
Los tratamientos combinados con múltiples moléculas pueden mejorar la eficacia clínica general de los fármacos anticancerosos actuales [68,106]. Debido a la resistencia a múltiples fármacos y la recurrencia del tumor, aún es urgente el desarrollo de nuevas estrategias para mejorar la sensibilidad a la quimioterapia y minimizar los efectos secundarios adversos. En este sentido, los flavonoides se han considerado candidatos prometedores en virtud de su actividad anticancerígena (Figura 5). Yuan y otros[107] proporcionó evidencia de la eficacia antiproliferativa de la combinación de arsenito y delfinidina (siendo este último uno de los compuestos de antocianina) en células NB4 y HL-60 APL humanas. Delfinidina sensibilizó a las células de leucemia resistentes al arsenito a la apoptosis modulando la cantidad de glutatión y reduciendo la actividad de NF-kB. También demostraron que el tratamiento combinado era selectivo, ya que aumentaba la citotoxicidad del arsenito contra las células cancerosas, pero no sobre las células mononucleares de sangre periférica humana [107].

Además, el tratamiento combinado con flavonoides ejerció efectos beneficiosos en varios tipos de células estabilizadas de tumores sólidos. Se ha demostrado que la quercetina sensibiliza las células de glioblastoma humano U87 y U251 a la temozolomida, un agente quimioterapéutico alquilante oral, in vitro a través de la inhibición de la proteína de choque térmico 27 [108]. Los flavonoides pueden ingresar al cerebro [109]. El potencial anticancerígeno de una combinación de isoflavonas biocanina A
y la temozolomida contra las células de glioblastoma U87 y T98G se asoció con una mayor expresión de p-p53, inhibición de la viabilidad celular y expresión de las proteínas de supervivencia celular EGFR, p-Akt, p-ERK, tipo membrana-MMP1 y c-myc[ 110]. El tratamiento combinado en células cancerosas indujo la detención del ciclo celular en la fase G1 y un cambio sustancial en el metabolismo energético de anaeróbico a aeróbico [95]. En las células de cáncer de colon, la fundición potenció la apoptosis inducida por el ligando inductor de apoptosis relacionado con TNF (TRAIL) a través de la regulación positiva del receptor de muerte 5 y la regulación negativa de las proteínas de supervivencia, como la survivina, Bdl-xL, Bcl-2, celular FLICE inhibidora similar a la proteína (cFLIP), y la proteína inhibidora de la apoptosis ligada al cromosoma X (XIAP)[95]. En células LoVo de adenocarcinoma colorrectal humano, Palko-Labuzet al. demostraron recientemente que el flavonoide baicaleína potencia el efecto antiproliferativo y proapoptótico de las estatinas, lo que hace que el tratamiento con doxorrubicina sea eficaz en una línea celular resistente [111]. Además, la catequina EGCG del té verde suprime el crecimiento tumoral y aumenta la eficacia terapéutica de los medicamentos en varios tipos de cáncer, como el 5-fluorouracilo (5-FU) en las células de cáncer de colon al inhibir la proteína regulada por glucosa 78 (GRP78)/NF-kB/miR-155-5p/MDR1 vía [112].
Se ha sugerido que el polifenol EGCG en el té tiene el potencial de ser un adyuvante terapéutico contra el cáncer de mama metastásico humano [113]. Un estudio clínico mostró que los pacientes con cáncer de mama sometidos a radioterapia y administración oral de EGCG presentaban una activación reducida de MMP-9/MMP-2 acompañada de niveles séricos bajos de VEGF y factor de crecimiento de hepatocitos (HGF)[113]. En una línea celular de cáncer de mama humano MDA-MB-231, la luteolina aumenta la acción de la doxorrubicina y el paclitaxel mediante la supresión de la señalización mediada por Nrf2-y el bloqueo de STAT3 [95,114]. Se observó una actividad similar para el flavonoide glabridina en líneas celulares de cáncer de mama resistentes a MDA-MB-231/MDR1 (con sobreexpresión de P-GP) y en células MCF-7/ADR (con sobreexpresión de P -GP y MRP2). El efecto sensibilizador de la glabridina puede deberse a su capacidad para aumentar la acumulación de doxorrubicina en las células MDA-MB-231/MDR1 mediante la supresión de la expresión de P-GP y la inhibición competitiva de la bomba de salida de P-GP, lo que mejora la actividad apoptótica de la doxorrubicina. [115]. Kundur et al. han demostrado que la quercetina y la curcumina administradas juntas tienen un efecto antitumoral sinérgico en las células de cáncer de mama triple negativo (TNBC), incluida la línea MDA-MB-231, mejorando la expresión de la proteína de susceptibilidad al cáncer de mama tipo 1 [116].
Recientemente, Moon y sus colegas informaron que el tratamiento con nobiletina aumentó la acumulación de adriamicina intracelular (ADR) en la línea celular humana NSCLC A549/ADR al promover la eficacia del tratamiento a través de un mecanismo acompañado de una regulación a la baja de la expresión de Akt, MYC (MYCN) derivado del neuroblastoma. ), GSK-3 , MRP1 y -catenina [117]. Además, en células de NSCLC resistentes a mutantes de EGFR, la apigenina junto con el inhibidor de la tirosina quinasa de EGFR gefitinib inhibieron factores oncogénicos importantes como c-Myc, el factor 1 alfa inducible por hipoxia (HIF-1a) y EGFR, y también redujo el uso de glucosa al suprimir la expresión de su transportador, lo que sugiere el posible uso de la combinación de las dos moléculas en la práctica clínica [118]. La activación de la vía de apoptosis intrínseca mediante la detención de la fase G1 y la expresión de fosfatasa aumentó la citotoxicidad de paclitaxel en células de cáncer de próstata tratadas con naringenina, un flavonoide polifenólico derivado de cítricos. Uno de los reguladores negativos clave de la vía de señalización de PI3K/Akt, el homólogo de tensina eliminado en el cromosoma 10 (PTEN), también está involucrado en este mecanismo, junto con la regulación a la baja de NF-kB, Snail, Twist y c-Myc. Expresión de ARNm y supresión de la migración celular [119]. Estos resultados sobre el uso combinado de las dos moléculas in vitro destacan su potencial terapéutico en el cáncer de próstata, aunque obviamente también es necesaria una evaluación detallada del mecanismo subyacente a la acción combinada in vivo.

3 Conclusiones
Los flavonoides han mostrado propiedades particularmente efectivas para contrarrestar el crecimiento tumoral y hacer que las células cancerosas sean resistentes a las terapias convencionales. Con la presente compilación de información de la literatura actual, se ha hecho un intento de resaltar el potencial de los flavonoides en la terapia del cáncer, ya sea que se usen solos o en combinación con agentes quimioterapéuticos. Aunque se ha destacado la eficacia potencial de los flavonoides para contrarrestar el crecimiento tumoral, la búsqueda de mecanismos de acción aún llevará mucho tiempo.
Contribuciones de autor: CF y SB concibieron la idea de escribir esta reseña. CF, MRIB, GF, GP, CT, CM y SB contribuyeron a la búsqueda y redacción de literatura. CF, SB, CM y CT revisaron el documento. C editó el documento. Todos los autores han leído y aceptado la versión publicada del manuscrito.
Fondos: Esta investigación no recibió financiación externa.
Expresiones de gratitud: MRand GPare destinatarios del Ph.D. Program in Evolutionary Biology and Ecology, Department of Biology, University of Rome Tor Vergata, Via Della Ricerca Scientifica, 00133 Rome, Italy). CT fue apoyado por Fondazione Umberto Veronesi, que se reconoce con gratitud.
Conflictos de interés: Los autores declaran no tener conflicto de intereses.
Referencias
1. Steck, SE; Murphy, EA Patrones dietéticos y riesgo de cáncer. Nat. Rev. Cáncer. 2020, 20, 125–138. [Referencia cruzada]
2. Marai, JPJ; Deavours, B.; Dixon, RA; Ferreira, D. La estereoquímica de los flavonoides. En La ciencia de los flavonoides; Springer: Nueva York, NY, EE. UU., 2007; págs. 1–35.
3. Panche, AN; Diwan, AD; Chandra, SR Flavonoides: una visión general. J. Nutr. ciencia 2016, 5, e47. [Referencia cruzada]
4. Middleton, E. Los flavonoides. Tendencias Pharmacol. ciencia 1984, 5, 335–338. 5. Xiong, Y.; Zhang, P.; Warner, DR; Fang, Z. 3-Colorante de desoxiantocianidina: naturaleza, salud, síntesis y aplicaciones alimentarias. compr. Rev. ciencia de los alimentos. Seguridad alimentaria 2019, 18, 1533–1549. [Referencia cruzada] [PubMed]
6. Khoo, ÉL; Azlán, A.; espiga, ST; Lim, SM Antocianidinas y antocianinas: pigmentos coloreados como alimentos, ingredientes farmacéuticos y los posibles beneficios para la salud. Nutrición alimentaria Res. 2017, 61, 1361779. [Referencia cruzada]
7. Hostetler, GL; Ralston, RA; Schwartz, SJ Flavonas: Fuentes alimenticias, biodisponibilidad, metabolismo y bioactividad. Adv. Nutrición 2017, 8, 423–435. [Referencia cruzada]
8. Aherne, SA; O'Brien, NM Flavonoles dietéticos: química, contenido de alimentos y metabolismo. Nutrición 2002, 18, 75–81. [Referencia cruzada]
9. Mazur, WM; Duque, JA; Wähälä, K.; Rasku, S.; Adlercreutz, H. Isoflflavonoides y lignanos en legumbres: aspectos nutricionales y de salud en humanos. Nutrición Bioquímica 1998, 9, 193–200. [Referencia cruzada]
10. Hammerstone, FJ; Lázaro, SA; Schmitz, HH Contenido de procianidina y variación en algunos alimentos de consumo común. J. Nutr. 2020, 130, 2086S–2092S. [Referencia cruzada]